For "konvergent evolusjon" tilbyr darwinister dårlig forklarende tukling
Av David Coppedge 30. januar 2026. Oversatt herfra. [Kursiv og understreking ved oversetter.}

Her er et dilemma for evolusjonister: Lignende designprinsipper og strategier finnes på tvers av riker. Planter og dyr bruker lignende proteiner som brytere for å aktivere immunresponsen når de invaderes av patogener. Historien fortelles i Science Magazine -lenke. Redaktørene kaller det "Delt logikk i ulike immunsystemer".
"Det medfødte immunforsvaret til både planter og dyr er avhengig av evnen til å gjenkjenne patogenavledede molekyler og stimulere en forsvarsrespons. Jones et al. gjennomgår hvordan denne felles funksjonen oppnås i så forskjellige riker av lignende molekyler. Gjenkjenningssystemet er bygget for følsomhet mot utløsere og konstruert på en modulær måte. å forstå slike funksjoner kan være nyttig for å bygge nye veier gjennom syntetisk biologi, enten det er for å utvide sykdomsforsvar eller konstruere nye signalresponskretser."
Standard darwinistiske termer
Forfatterne, Jones, Vance og Dangl, bruker standard darwinistiske termer gjennomgående - felles stamfar, naturlig utvalg, samevolusjon osv. Likevel kan de ikke unngå å legge merke til hvor lik "strategien" er i "evolusjonært gamle" slektslinjer. Siden mekanismene i stor grad er "konservert" mellom planter og dyr, tilskriver de det den vanlige unnvikende, sirkulære forklaringen: konvergent evolusjon.
Bilde 1. Arabidopsis-planten
"Flercellulære eukaryoter samevolverer med mikrobielle patogener, som utøver sterkt selektivt press på immunforsvaret til vertene sine. Planter og dyr bruker intracellulære proteiner fra Nukleotidbindende domene, Leucin-Rike repetisjoner (NLR) superfamilien for å oppdage mange typer mikrobielle patogener. NLR-domenearkitekturen utviklet seg sannsynligvis uavhengig og konvergent i hvert rike, og de molekylære mekanismene for patogendeteksjon av plante- og dyre-NLR-er har lenge blitt ansett som forskjellige. Imidlertid overlapper mikrobielle gjenkjenningsmekanismer hverandre, og det er nå mulig å skille ut viktige sentrale trans-rike-prinsipper for NLR-avhengig immunfunksjon. Her forsøker vi å formulere disse prinsippene."
Deres grunnleggende "prinsipp"går slik: Hvis noe fungerer, må det ha utviklet seg.
"Vi foreslår at NLR-arkitekturen har utviklet seg for patogen-registrering i ulike organismer på grunn av dens nytteverdi som en tett foldet "håravtrekker"-enhet der et praktisk talt ubegrenset antall mikrobielle deteksjonsplattformer kan integreres."
Hvor smart av separate riker av organismer å ha funnet ut av dette, uavhengig av hverandre! Forfatterne bare vet at disse tingene utviklet seg, for som Phillip Johnson en gang sa: "Hvis vitenskapen i det hele tatt skal ha noen forklaring på biologisk kompleksitet, må den nøye seg med det som er igjen når det uakseptable er utelukket" (Darwin on Trial, 1991, s. 28). Vi ser den vanskelige forklarende tuklingen i antall ganger forfatterne bruker ordet "sannsynligvis" assosiert med "utviklet seg":
"NLR-domenearkitekturen utviklet seg sannsynligvis uavhengig og konvergent i hvert rike…."
"..Drosophila DARK og nematoden CED4, og utviklet seg sannsynligvis fra en klasse prokaryote ATPaser.."
"Dermed utviklet plante- og dyre-NLR-er seg sannsynligvis fra distinkte forfedre-NBD-linjer.."
"Dette antyder at paraloger av ekte mål for virulensproteiner kan ha utviklet seg til å ligne det målet.."
"NLR-er sannsynligvis avledet fra en felles forfader.."
"Animalske NLR-er, derimot, bærer en distinkt NBD-subtype .. som også sannsynligvis er avledet fra et distinkt prokaryot forfedredomene."
 Med all konkurranse eliminert
Med all konkurranse eliminert
Argumentasjon gjennom påstander er enkelt når all konkurranse er eliminert fra feltet. Men de stusser på det:
"Til tross for bemerkelsesverdig mangfold i oppstrøms og nedstrøms signalhendelser, er vi tvunget til å tenke over hva som er så fundamentalt fordelaktig med NLR-arkitekturen som kan forklare hvorfor den oppsto konvergent i planter og dyr for å spille en rolle i patogendeteksjon og forsvarsaktivering."
Det er sant at det er forskjeller i antall NLR-proteiner og deres nedstrøms aktiveringer, men begge kongedømmene ser ut til å bruke fire lignende strategier, hvorav minst tre er felles for både planter og dyr. Og det kan være enda mer fellestrekk:
"Hele spekteret av mekanismer i hvert kongedømme antyder at det er rom for flere konseptuelle likheter enn tidligere antatt. Gitt dette mangfoldet foreslår vi at en fordel med NLR-arkitekturen ganske enkelt kan være dens evne til å fungere som en robust av/på-bryter i ulike signalkontekster."
Figur 2 viser et "NLR-o-gram" mellom menneskelige gener og gener fra laboratorieplanten Arabidopsis (bildet øverst). Likhetene er slående til tross for den lange evolusjonstiden etter diversifisering fra en encellet organisme, som utvilsomt manglet alle de koordinerte delene av systemet.
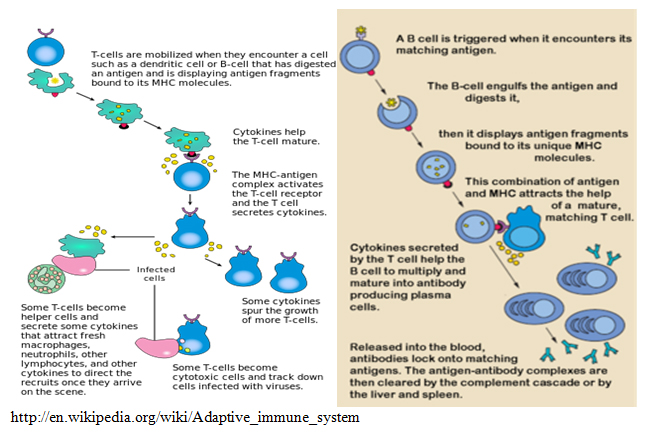
Bilde 2. Eks. Hvordan immunforsvar benytter T-celler
Kanskje NLR-er lett kan drive fra organisme til organisme? Det ser ikke ut til å være tilfelle:
"Funksjonell overføring av plante-NLR-er på tvers av artsbarrierer har vist seg å være stort sett umulig. Denne begrensede taksonomiske funksjonaliteten er dårlig forstått…."
Det er ikke det eneste som er dårlig forstått. Kompleksiteten i NLR-mekanismer lukker døren for unnskyldningen om at de er enkle og lett å utvikle:
"Til tross for gjennombrudd i vår molekylære forståelse av NLR-aktivering, gjenstår mange viktige spørsmål. Biokjemiske mekanismer for NLR-aktivering forblir uklare. Hendelser nedstrøms for plante-NLR-aktivering og utganger som transkripsjon av forsvarsgener, endringer i cellepermeabilitet, lokalisert celledød og systemisk signalering forblir  uklare. Vi vet ikke om aktiverte plante-NLR-er oligomeriserer eller, hvis de gjør det, hvordan dette oppnås, gitt mangfoldet av subcellulære aktiveringssteder observert for forskjellige NLR-er. Det er ikke klart om og hvordan de forskjellige N-terminale domenene til plante-NLR-er signaliserer. Vi har økende kunnskap om hvordan dyre-NLR-er setter seg sammen og signaliserer, selv om det fortsatt er kunnskapshull."
uklare. Vi vet ikke om aktiverte plante-NLR-er oligomeriserer eller, hvis de gjør det, hvordan dette oppnås, gitt mangfoldet av subcellulære aktiveringssteder observert for forskjellige NLR-er. Det er ikke klart om og hvordan de forskjellige N-terminale domenene til plante-NLR-er signaliserer. Vi har økende kunnskap om hvordan dyre-NLR-er setter seg sammen og signaliserer, selv om det fortsatt er kunnskapshull."
Oppskalering av situasjonen
Vi finner et komplekst system som involverer hårfine utløserbrytere laget av proteiner. Disse NLR-proteinene samhandler med mange andre komponenter i cellen. I hver art er de forskjellige nok til å forhindre overføring mellom arter, men de deler felles funksjoner og bruker lignende strategier.
Høres ikke dette litt ut som bildeler? Når du går til delebutikken, kjøper du en lyddemper eller tennplugger eller et luftfilter som passer til kjøretøyet ditt. Fyren bak pulten slår opp merke og modell, og finner modellnummeret som passer for deg. Det er åpenbare likheter i funksjon, men du setter ikke et luftfilter fra gressklipperen din inn i en pickup.
Du kan se en viss "evolusjon" av luftfiltre mellom forskjellige modellår av samme lastebil, men vi vet alle at hver del er intelligent tilpasset kjøretøyet sitt, fordi delene må fungere sammen.
Bilde 3. Menneskets immunforsvar deler proteiner og funksjon med andre rikers immunforsvar
Hvorfor ikke vurdere design?
Siden evolusjonistene er forvirret av sin egen forklaring, ønsker vi å frigjøre dem fra tvangstrøyen Darwin har satt dem i. I stedet for å se på immunresponsen som et nytt mystisk eksempel på "konvergent evolusjon", hvorfor ikke vurdere intelligent design av "prinsippene" og "strategien" som brukes av forskjellige typer organismer? Og når det ser ut til å være noen fellestrekk mellom arter, slekter og familier, hvorfor ikke vurdere mulige designprinsipper som er iboende i systemet - en slags forhåndsprogrammert robusthet - for å tillate modifikasjoner etter hvert som forholdene endrer seg?
Vi prøver bare å hjelpe, siden dere innrømmet å være "tvunget til å tenke over" fordelene med "arkitekturen" dere har undersøkt. å tenke på design kan også gjøre det lettere å nå målet som er nevnt øverst: "å konstruere nye signalresponskretser". Konstruksjon av kretser gjøres vanligvis best med intelligent design.
(Opprinnelig publisert i 2016)
 Bilde 4: Om forfatteren: DAVID COPPEDGE -kredit David Coppedge.
David Coppedge er en frilans vitenskapsreporter i Sør-California. Han har vært styremedlem i Illustra Media siden grunnleggelsen og fungerer som deres vitenskapskonsulent. Han jobbet ved NASAs Jet Propulsion Laboratory (JPL) i 14 år, på Cassini-oppdraget til Saturn, til han ble kastet ut i 2011 for å dele materiale om intelligent design, en diskriminerende handling som førte til en nasjonalt publisert rettssak i 2012. Discovery Institute støttet saken hans, men en ensom dommer dømte ham mot ham, uten forklaring. En naturfotograf, friluftsmann og musiker, David har B.S. grader i realfagsutdanning og i fysikk og holder presentasjoner om ID og andre vitenskapelige emner.
Bilde 4: Om forfatteren: DAVID COPPEDGE -kredit David Coppedge.
David Coppedge er en frilans vitenskapsreporter i Sør-California. Han har vært styremedlem i Illustra Media siden grunnleggelsen og fungerer som deres vitenskapskonsulent. Han jobbet ved NASAs Jet Propulsion Laboratory (JPL) i 14 år, på Cassini-oppdraget til Saturn, til han ble kastet ut i 2011 for å dele materiale om intelligent design, en diskriminerende handling som førte til en nasjonalt publisert rettssak i 2012. Discovery Institute støttet saken hans, men en ensom dommer dømte ham mot ham, uten forklaring. En naturfotograf, friluftsmann og musiker, David har B.S. grader i realfagsutdanning og i fysikk og holder presentasjoner om ID og andre vitenskapelige emner.
Oversettelse, via google oversetter, og bilder ved Asbjørn E. Lund